2.3. Le message nerveux
2.3.1. Synapses
Pour comprendre comment le message nerveux est acheminť jusqu'au cerveau, il est impťratif de comprendre comment celui-ci est transmis d'une cellule nerveuse ŗ une autre. Il nous faut donc expliquer le fonctionnement d'une synapse contactant les deux cellules : l'extrťmitť du prolongement prť-synaptique est formť d'un renflement, le bouton synaptique, riche en neurotransmetteur, contenu dans de petites vťsicules. Un espace sťpare le neurite prť-synaptique du neurite post-synaptique appelť la fente synaptique. La membrane post-synaptique (qui doit recevoir l'influx) porte des rťcepteurs spťcifiques ŗ ces neurotransmetteurs ; et lorsqu'un un influx nerveux atteint le bouton synaptique, il y provoque l'expulsion du neuromťdiateur dans la fente par ťclatement des vťsicules : celui ci atteint les sites rťcepteurs de la membrane post-synaptique et y dťclenche un influx nerveux. Dans le cas ťtudiť, pour que le neurotransmetteur, en l'occurrence le glutamate, soit libťrť par les cŰnes, c'est ŗ dire pour que les vťsicules ťclatent, il faut que le potentiel de rťcepteur ŗ proximitť de la synapse se situe ŗ un certain seuil de dťpolarisation ; la quantitť libťrťe augmentera ensuite en fonction du niveau de dťpolarisation.
Figure 2-6. La transmission synaptique

Lťgende.
Stockage du glutamate dans une vťsicule synaptique;
L'arrivťe du potentiel de rťcepteur dans la terminaison prťsynaptique;
Fusion des vťsicules avec la membrane prť-synaptique;
Libťration du glutamate dans la fente synaptique;
Fixation du glutamate sur les rťcepteurs de la membrane postsynaptique;
Nouveau potentiel de rťcepteur;
Inactivation du glutamate par des enzymes;
Recapture du glutamate.
Dans l'obscuritť, comme nous l'avons expliquť, par suite du courant d'obscuritť, et de leur potentiel de rťcepteur de -40mV, les cŰnes sont en permanence dťpolarisťs; leur neurotransmetteur, le glutamate, est donc libťrť en continu. Par contre, lors de la stimulation lumineuse, le potentiel de rťcepteur des cŰnes s'accroÓt (passant de -40mV ŗ -80mV), s'ťloignant ainsi brusquement du seuil de dťpolarisation nťcessaire ŗ la libťration complŤte du glutamate : il y aura ainsi moins de glutamate libťrť.
Ainsi, le potentiel de rťcepteur, contenant les informations sensorielles nťes lors de la transduction, contrŰle le flux du neurotransmetteur libťrť comme nous venons de le dťcrire. Le glutamate libťrť, lui, se chargera d'induire les informations sensorielles dans les cellules bipolaires et horizontales ; nous allons maintenant ťtudier les "transmissions synaptiques" entre les cŰnes et les cellules bipolaires dans l'obscuritť et lors de la stimulation lumineuse.
2.3.2. CŰnes/Bipolaires
Comme nous l'avons vu, les cŰnes prťsentent deux types de synapses. Chacune contacte deux catťgories de cellules bipolaires : les bipolaires invaginťes (BipI) et les bipolaires superficielles (BipS), les zones de contact se situant dans la couche plexiforme externe. Nous avons ťgalement vu que l'activation de la iodopsine par la lumiŤre se traduisait au niveau synaptique par une moindre quantitť de neurotransmetteur libťrť. Inversement, sitŰt l'activation terminťe, cette quantitť augmente. Le point capital ici est de comprendre que ces deux catťgories de cellules bipolaires rťagissent de faÁon opposťe au glutamate, le neurotransmetteur libťrť par le cŰne. Ceci sous-entend qu'il existe deux types de synapses dont le rŰle diffŤre: les synapses excitatrices et les synapses inhibitrices. En effet, la synapse peut soit transmettre une information, soit l'empÍcher de se propager. Dans la synapse excitatrice, la libťration du neurotransmetteur provoque du cŰtť post-synaptique une augmentation de la permťabilitť au sodium Na+. Du sodium Na+ pťnťtrant dans le neurone post-synaptique, la membrane de celui ci est dťpolarisťe, et un potentiel post-synaptique excitateur apparaÓt. L'information passe d'un neurone ŗ l'autre. Dans la synapse inhibitrice, la libťration du neurotransmetteur entraÓne au niveau du neurone post-synaptique une hyperpolarisation de la membrane. Celle-ci est donc moins sensible aux stimulations et on assiste ŗ une inhibition de la transmission de l'influx. Ainsi:
dans l'obscuritť, sous l'action du glutamate constamment libťrť, les bipolaires invaginťes sont hyperpolarisťes et les superficielles dťpolarisťes. Car, les synapses des BipI ťtant inhibitrices, la libťration du glutamate entraÓne une hyperpolarisation de leur membrane; de la mÍme maniŤre, dans les synapses excitatrices des BipS, la libťration du neuromťdiateur provoque la dťpolarisation et donc l'excitation de leur membrane, avec apparition d'un nouveau potentiel membranaire, contenant l'information visuelle du potentiel de rťcepteur.
au dťbut de la stimulation lumineuse, avec donc moins de glutamate libťrť, l'hyperpolarisation des bipolaires invaginťes se trouve rťduite ; cette variation se fait donc dans le sens d'une dťpolarisation et donc d'une excitation; un potentiel membranaire apparaÓt, et contient l'information visuelle du potentiel de rťcepteur. En revanche, pour les bipolaires superficielles, la variation se fait en sens contraire, en direction d'une hyperpolarisation.
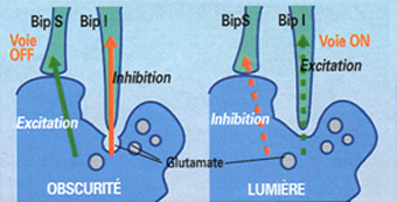
On remarque ainsi que le dťbut et la fin de la stimulation lumineuse sont caractťrisťs par des dťpolarisations successives des deux catťgories de cellules bipolaires de cŰnes : les bipolaires invaginťes au dťbut, puis les bipolaires superficielles ŗ la fin. Le point essentiel ŗ retenir est que, dŤs cet ťtage prťcoce du traitement rťtinien, il existe deux voies complŤtement indťpendantes qui mesurent, par des excitations, l'une l'augmentation de la quantitť de lumiŤre (BipI), l'autre sa diminution (BipS). Ces deux voies, respectivement ON et OFF, fonctionnent en parallŤle et demeurent indťpendantes jusqu'au niveau du cortex cťrťbral primaire.
2.3.3. Bipolaires/Ganglionnaires
Mais avant de quitter l'oeil, un dernier ťtage doit Ítre franchi par l'information visuelle : celui des cellules ganglionnaires. Entre elles et les bipolaires des cŰnes, le glutamate sert encore de neurotransmetteur. Mais cette fois, son action va toujours dans le mÍme sens : dťpolarisation, et donc excitation. La cellule ganglionnaire sera excitťe quand la cellule bipolaire qui la contacte le sera aussi. Suivant le principe des deux voies, ON et OFF, les ganglionnaires ON seront les cellules connectťes par les bipolaires ON (BipI) et les ganglionnaires OFF celles connectťes par les bipolaires OFF (BipS). Le glutamate se chargera d'induire ŗ nouveau les informations visuelles contenues dans les potentiels membranaires dans les cellules ganglionnaires, entraÓnant une fois de plus l'apparition d'un potentiel membranaire, contenant l'information sensorielle initiale, dťjŗ traitťe par les bipolaires. Les contacts entre les cellules bipolaires et les ganglionnaires ont lieu dans une couche de la rťtine appelťe couche plexiforme interne. Or, les connexions entre les BipI et les ganglionnaires ON d'une part, les BipS et les ganglionnaires OFF d'autre part, s'effectuent dans des sous-couches distinctes et superposťes de cette couche. Tout se passe donc comme si cette derniŤre etait constituťe de deux reprťsentations antagonistes de la mÍme image optique, l'une ťtant le nťgatif de l'autre. Les liaisons cŰnes-cellules bipolaires-cellules ganglionnaires forment les voies de transmission directe. Il existe aussi des voies parallŤles qui comprennent les cellules horizontales mettant en liaison diffťrents types de cŰnes et les cellules amacrines mettant en liaison diffťrentes cellules bipolaires.
Les cellules ganglionnaires possŤdent une longue fibre appelťe axone. Nous avons dťjŗ vu que l'ensemble des axones constituait le nerf optique. Pour pouvoir crťer des signaux capables d'atteindre le cerveau sans perte d'information, il faut que les potentiels d'action propagťs soient gťnťrťs au niveau de ces fibres optiques ; pour cela, la dťpolarisation de la membrane de l'axone est indispensable (valeur seuil). Celle ci provoquera donc l'apparition d'un potentiel d'action (PA). Ce PA est en fait une inversion brutale et transitoire du potentiel membranaire, qui obťit a la loi du tout ou rien ; c'est ŗ dire que si le seuil de dťpolarisation n'est pas atteint, il n'apparaÓt pas ; mais si le seuil est atteint, la rťponse est maximale d'emblťe. Le PA se propage aussi sans attťnuation, de maniŤre autonome, tout au long de la fibre de l'axone. Les ganglionnaires ON envoient donc des influx le long des fibres optiques au dťbut de la stimulation lumineuse; les ganglionnaires OFF ŗ la fin. Le message nerveux en amplitude est transformť en un message codť par la frťquence des potentiels d'action. Le nerf optique les transmet finalement ŗ la zone optique du cerveau. A ce dernier la tache de dťcoder, ťtape par ťtape, les potentiels d'actions, pour reformer une image en trois dimensions et en couleurs. Cette dichotomie ON/OFF signifie que l'image rťtinienne, ťchantillonnťe par les cŰnes, est subdivisťe en deux sous-images. L'une est formťe par les augmentations locales de la quantitť de lumiŤre, l'autre par ses diminutions.
Ainsi, nous avons vu qu'avant d'Ítre transmise par les fibres du nerf optique, l'image est traitťe par plusieurs cellules nerveuses. En effet, les cŰnes se chargent de mesurer, point par point, sur l'image oculaire, le contenu ťnergťtique de la lumiŤre qu'ils captent et de traduire ces mesures par l'amplitude du potentiel rťcepteur. Les autres cellules rťtiniennes se chargent de collecter et d'intťgrer des signaux ťchantillonnťs par un certain nombre de cŰnes, distribuťs sur des surfaces plus ou moins ťtendues de la mosaÔque rťtinienne. Ces surfaces, ces zones de collecte s'appellent des champs rťcepteurs.
2.3.4. Champs rťcepteurs
Ceux-ci ont une forme grossiŤrement circulaire (de 100 μm de diamŤtre), pouvant Ítre divisťe en deux zones concentriques. La figure ci dessous montre l'organisation de ces champs rťcepteurs rťtiniens.
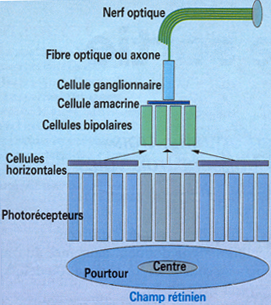
L'ensemble des photorťcepteurs en relation avec une mÍme cellule ganglionnaire forme son champ rťcepteur. Au centre, la population de cŰnes en contact synaptique direct. A la pťriphťrie, la population de cŰnes connectťe ŗ la cellule bipolaire via les cellules horizontales.
Celles-ci possŤdent la caractťristique d'inverser les signaux ťmis par les photorťcepteurs de la pťriphťrie; ainsi, s'ils sont soumis aux mÍmes conditions d'ťclairement, les cŰnes des deux zones (centre et pourtour) exercent des influences antagonistes sur la cellule bipolaire. Prenons l'exemple d'une cellule bipolaire ON (BipI). Nous avons vu qu'un supplťment de lumiŤre dans le centre de son champ rťcepteur l'excitait. Simultanťment, un supplťment de lumiŤre de son pourtour va l'inhiber. Ainsi, quand centre et pourtour sont soumis ŗ la mÍme variation de lumiŤre, l'excitation par le centre et l'inhibition par le pourtour se contrecarrent (les signaux s'annulent), et la cellule bipolaire reste "muette". Par contre, la mÍme cellule rťpond de faÁon optimale quand le centre est illuminť et, en mÍme temps, le pourtour assombri. Inversement, dans le cas d'une bipolaire OFF (BipS), celle ci rťpondra de faÁon optimale quand le centre de son champ rťcepteur sera sombre et son pourtour ťclairť. En d'autres termes, la cellule bipolaire mesure le contraste de lumiŤre entre le centre et le pourtour de son champ rťcepteur.

Etant donnť que l'activitť des trois catťgories de cŰnes, S, M et L, est restreinte ŗ trois rťgions distinctes du spectre, le bleu, le vert, et le rouge, le contraste mesurť pourra porter sur des comparaisons chromatiques entre des populations distinctes de cŰnes. Selon le type d'articulations assurť par les cellules horizontales entre ces cŰnes, trois sortes d'opposition existent:

les signaux des cŰnes M s'opposent ŗ ceux des cŰnes L: vert/rouge
les signaux des cŰnes S ŗ la somme des signaux issus des cŰnes M et L (correspondant au jaune): jaune/bleu
enfin, quand les cellules horizontales mťlangent les signaux des diffťrentes catťgories de cŰnes, l'opposition est achromatique et porte seulement sur la diffťrence entre le clair et le sombre.
Ainsi, certaines couleurs se mťlangent mieux que d'autres. Par exemple, il est difficile d'imaginer un vert rouge‚tre ou du jaune bleu‚tre, des couleurs opposťes. Bien entendu, la distinction ON et OFF porte sur chacune de ses voies.
Nous venons ainsi d'ťtudier le cas des cŰnes situťs dans la zone pťriphťrique de la rťtine, or il y a aussi des cŰnes dans la fovťa, qui sont d'ailleurs, comme nous l'avons dťjŗ dit, beaucoup plus nombreux. Les cŰnes de la fovťa sont elles en relation avec une seule cellule bipolaire, elle-mÍme en relation avec une seule cellule ganglionnaire. Il y a donc, pour chaque cŰne une fibre nerveuse spťcifique (axone de la cellule ganglionnaire): cela explique la vision trŤs fine que procure la fovťa. Cellules horizontales et cellules amacrines interviennent en tant que modulateurs sur cette chaÓne, comme dans le cas des cŰnes de la pťriphťrie.